
Hieu Huu Le 1,2,* , Majid Shakeri 1, Hafiz Ansar Rasul Suleria 1 , Weicheng Zhao 1, Rachel Mai McQuade 3, Deborah Jayne Phillips 1, Eva Vidacs 1, John Barton Furness 1,4 , Frank Rowland Dunshea 1,5 , Valeria Artuso-Ponte 6 and Jeremy James Cottrell 1,*
1 墨尔本大学兽医和农业科学学院,维多利亚州帕克维尔,3010,澳大利亚;
2 国立农业大学动物科学学院,越南特劳奎,嘉林,河内131004, 越南
3 墨尔本大学弗洛里神经科学与心理健康研究所,维多利亚州帕克维尔,3010,澳大利亚;
4 墨尔本大学解剖学和神经科学系,维多利亚州帕克维尔,3010,澳大利亚
5 利兹大学生物科学学院,利兹LS2 9JT,英国
6 仹犇泰 Phytobiotics Futterzusatzstoffe GmbH, D-65343 埃尔特维尔, 德国;
* 通信:huul1@student.unimelb.edu.au(H.H.L.);jcottrell@unimelb.edu.au
摘要:
热应激(HS)影响肉猪的生产力,部分原因是氧化应激和炎症反应增加导致的,特别是在胃肠道内的氧化应激和炎症反应。本研究旨在探讨植物来源的甜菜碱和异喹啉生物碱是否可以改善猪的热应激(HS)条件下的各项性能。50头雌性大白×长白长白种猪,适应对照组(CON),添加甜菜碱(BET)或添加异喹啉生物碱(IQA)日粮14天,然后暴露于热应激或热中性条件。甜菜碱(BET)和异喹啉类生物碱(IQA)均部分改善了与热应激(HS)疾病相关的呼吸频率(p = 0.013)和直肠温度(p = 0.001)的升高。热应激反应增加唾液皮质醇浓度,降低血浆肌酐、乳酸和甲状腺激素浓度。热应激反应增加了结肠FD4通透性,异喹啉类生物碱(IQA)降低了结肠FD4通透性(p = 0.030)。热应激增加空肠和回肠的炎症,如空肠白细胞介素-1β (p = 0.022) 升高以及回肠白细胞介素-1β (p = 0.004) 和白细胞介素-8 (p = 0.001) 升高所示。热应激(HS)组在血浆总抗氧化能力(TAC)方面没有观察到差异,但与异喹啉类生物碱(IQA)相比,甜菜碱增加了血浆TAC。日粮甜菜碱(BET)增加了空肠,回肠(p < 0.001),血浆,肝脏,肾脏(p < 0.010),尿液(p = 0.002)中的甜菜碱浓度,并且肌肉中往往更高(p = 0.084)。甜菜碱浓度不受热应激(HS)的影响,但在血浆中往往较高并在肝脏中蓄积。这些数据表明,添加甜菜碱和异喹啉生物碱可改善生长猪热应激的后果,并防止热应激(HS)引起的结肠通透性增加。
关键词:热应激;生物 碱;甜菜 碱;抗 氧化剂;渗透性;肠道健康;温度调节
1. 简介
猪对热应激(HS)敏感,因为它们缺乏功能性汗腺,无法通过皮肤蒸发途径促进热量散发。此外,生长和胴体性状遗传选择的改善导致代谢产热增加,从而降低了猪的耐热能力。热应激影响了肉猪的高效生产,部分原因是采食量减少。然而,其他物种的配对喂养研究表明,仅靠采食量减少就可以解释生产力损失的一半[1]。生产力进一步下降的原因多方面的,但包括胃肠道(GIT)的破坏等因素。这是因为高热期间增加环境热损失的主要体温调节机制是将血流从内脏器官重新分配到唿吸道和皮肤[2-4]。由此导致的胃肠道(GIT)血流量减少不足以满足代谢需求,并导致氧化应激和绒毛上皮丢失,暴露固有层[5,6]。这导致内毒素和炎症的通透性增加[7,8]。
随着夏季热浪的发生率和严重程度的增加,人们开始关注开发改进的改善策略来对抗热应激,其中一种方法是通过优化营养[9]。也许最广为人知的添加剂是有机渗透剂——甜菜碱,它具有许多抵消热应激(HS)影响的特性。其中包括渗透液甜菜碱会降低膜结合ATP酶的活性[10],这可能有助于降低热应激动物的基础代谢率和直肠温度[11,12]。
作为甲基供体的其他特性可能有助于广泛的生物反应,减少氧化应激和甲基化途径,提高热应激牲畜的乳制品[13],肉类[14]和鸡蛋生产[15]的效率。植物来源的异喹啉生物碱(如血根碱)可减少运输猪的应激和沙门氏菌脱落[16,17]。异喹啉生物碱抑制细胞Na+/K+ATP酶,后者是产热的重要贡献者[18,19],因此可能提供缓解热应激的方法。此外,异喹啉生物碱具有抗氧化[20]和抗炎特性[20,21],可以抵消热应激(HS)期间普遍存在的氧化应激,炎症和胃肠道(GIT)粘膜破坏。因此,本实验旨在研究添加异喹啉生物碱和甜菜碱是否可以改善体温调节反应并保护经历热应激(HS)的生长猪的肠道完整性。
2. 材料和方法
2.1. 动物和实验设计
所有实验程序均由澳大利亚墨尔本大学兽医与农业科学学院动物伦理委员会批准(协议号1814434.1),并遵循澳大利亚科学目的动物护理和使用守则(第8版)(国家健康与医学研究委员会,2013年)的规定。
共有50头雌性养殖猪(大白×长品种,27.3±1.7公斤,平均±标准偏差)被随机分配到三种实验日粮处理组:对照(CON,n = 18,标准种植者日粮),标准日粮添加1克甜菜碱/公斤饲料(BET,n = 16)或标准日粮与0.15克/千克异喹啉生物碱提取物(Sangrovit® Extra, 仹犇泰Phytobiotics Futterzusatzstoffe GmbH,德国埃尔特维尔)/千克饲料(IQA,n = 16)。 Sangrovit® Extra是由博落回Macleaya cordata制备的提取物,含有异喹啉生物碱的混合物。先前已经表明,博落回提取物Macleaya cordata果荚中存在的主要生物碱(mg/g干物质)是血根碱(32.08),白屈菜红碱(7.36)和二氢血根碱(3.14),而整株在地上部分发现的主要生物碱是原阿片碱(7.93),别隐品碱(6.77)和血根碱(4.51)[21]。此外,博落回Macleaya cordata含有白屈菜红碱(1.1)和其它微量的血桂氨酸、高胆子多宁、去甲血根碱、氧血根碱和氧胆红碱[21]。猪在热中性(TN)条件下适应日粮14天(20°C;35-50%相对湿度)。基于小麦和菜籽粕的对照组(CON)日粮是为了满足国家研究委员会(2012)建议的营养需求而制定的(表1)。对照组(CON)日粮中每公斤含有13.4 MJ的ME和19.0%的粗蛋白。异喹啉类生物碱(IQA)日粮含有饲料添加剂制剂,可提供约1-1.5%的异喹啉类生物碱(IQA)。在适应期之后,每种日粮的一半猪要么继续在热中性(TN)条件下饲养,要么在“热应激”(HS)条件下饲养,其中包括8小时/天35°C(0900至1700小时)和28°C16小时/天(1700至0900小时)3天(TN和热应激(HS)的n = 25)。为了消除不同采食量的影响,所有猪均以ME×2.5倍(约占随意采食量的75%)配对饲喂[22],并提供自由饮水。
表 1.基础日粮的成分和组成
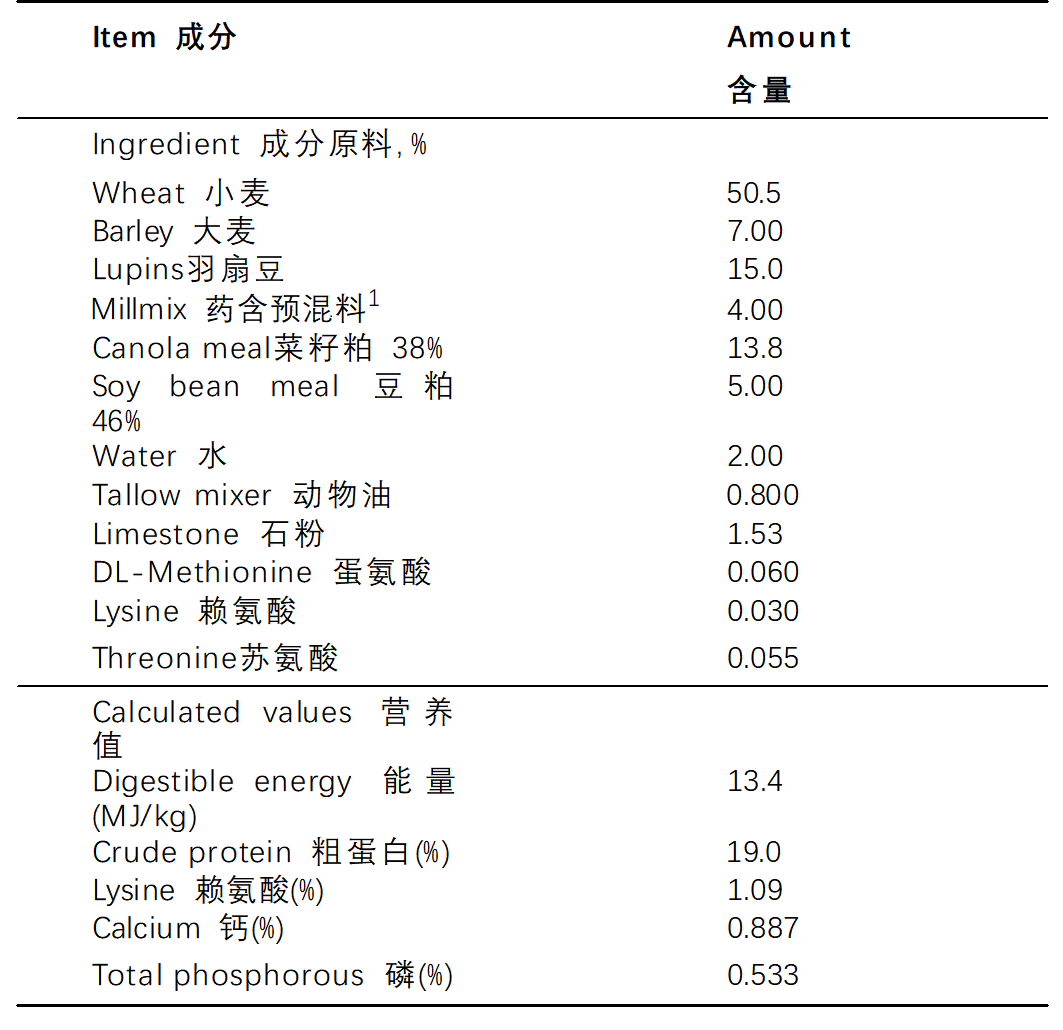
1 调整了Millmix的量,在日粮中分别以0.15和1 g / kg的含量添加Sangrovit Extra和甜菜碱。通过测定确认测试日粮中血根碱的最终浓度为0.61mg / kg(第2.10节)。
2.2. 生理观察
在环境挑战的第1天和第2天,通过每2 h对猪的唿吸速率(RR)、直肠和皮肤温度(分别为RT和ST)进行每日监测,评估猪的热应激。通过使用秒表计算侧翼运动20秒来观察唿吸速率(RR),然后以唿吸/分钟表示。使用插入直肠约2厘米的数字温度计(Surgipack,澳大利亚)测量直肠温度,直到温度稳定并使用手持式红外温度计(货号QM7221,非接触式红外温度计,Digitech)记录侧面的皮肤温度。
2.3. 唾液收集和唾液皮质醇测量
唾液在环境挑战的第1天和第2天通过拭子(Salivette® Cortisol,Sarstedt AG&Co,Nümbrecht,德国)在1500小时收集。 然后将拭子离心(室温下1000×g)2分钟以获得唾液,唾液储存在-80°C直至分析。唾液样品中的皮质醇浓度根据制造商的说明,使用商业试剂盒通过酶联免疫吸附测定法 (ELISA) 进行测量(Cat no. 1-3002-5,Salimetrics LLC,州立大学,宾夕法尼亚州,美国)。
2.4. 血液和组织采集、血气测量和安乐死
在环境挑战的第3天,猪被注射氯胺酮(20毫克/千克)和甲苯噻嗪(3毫克/千克)麻醉。麻醉后,从耳静脉收集 1 mL 新鲜血液样本并转移到血气分析仪 (EPOC®;美国马萨诸塞州沃尔瑟姆市阿莱尔)。然后使用含有EDTA抗凝剂的真空容器从颈外静脉收集第二份血液样本(5mL)(BD澳大利亚,北莱德,新南威尔士州,澳大利亚)。在4°C下以1500×g离心10分钟后收集血浆。将血浆样品储存在-20°C直至分析。
2.5. 血浆激素和代谢物测量
血浆游离三碘甲状腺原氨酸(T3)和甲状腺素(T4)按照制造商的说明(货号07-221105,MP生物医学,奥兰治堡,纽约,美国)用放射免疫测定试剂盒进行测量。T3的灵敏度和测定内CV分别为0.06 pg/dL和5.12%,T4的灵敏度和测定内CV分别为0.45 pg/dL和5.12%。血浆中的总抗氧化能力(TAC)是根据制造商的说明使用商业试剂盒(Cat No. 709001,开曼化学,密歇根州安娜堡,美国)测量的。结果表示为Trolox等效抗氧化能力,平均测定间和测定内CV分别为2.04%和1.47%。同样,血浆白蛋白、甘油三酯、胆红素浓度根据制造商的说明,使用981767、981786、981897(Thermo-Fisher,美国马萨诸塞州沃尔瑟姆)提供的试剂进行定量。白蛋白的测定间和测定内CV分别为9.4%和11.9%,甘油三酯为3.8%和2.1%,胆红素为4.9%和2.0%。
2.6. 尿液分析
安乐死后不久,使用透壁针和注射器直接从膀胱收集尿液样本。立即测量pH值(Eutech pH 5+ pH计,Thermo Fisher Scientific,美国马萨诸塞州沃尔瑟姆),并量化渗透压浓度(Advanced Micro Osfusion ometer 3300,Advanced Instruments,美国马萨诸塞州诺伍德)。根据制造商的说明,使用商业试剂(Cat No. 981767, 981897,Thermo Fisher Scientific,美国马萨诸塞州沃尔瑟姆)定量尿白蛋白和胆红素浓度,白蛋白测定间和测定内变化分别为7.5%和0.9%,胆红素为8.4%和2.7%。根据制造商的说明(第 500701 类,Cayman Chemical,开曼化学,美国,使用商业试剂盒定量尿肌酐浓度,测定间和测定内 CV% 分别为 3.1% 和 1.0%。
2.7. 组织取样和肠通透性测量
安乐死后立即收集回肠远端(~30 cm回盲部近端)和近端结肠(紧邻回盲部交界处远端)切片,并置于冷冻磷酸盐缓冲盐水中(0.9%氯化钠静脉注射,Baxter Healthcare,Old Toongabbie,新南威尔士州,澳大利亚),然后转移到克雷布斯溶液。通过钝性解剖去除黏膜并安装在圆形滑块上,并放入由两部分组成的Ussing室(EasyMount扩散室,生理仪器,美国加利福尼亚州圣地亚哥)中,如[23]所述。之后,将200 μL 25 mg/mL荧光素异硫氰酸酯-葡聚糖(FD4)(Sigma-Aldrich,圣路易斯,密苏里州,美国)加入腔室的粘膜侧,并在0、60和120分钟从腔室浆膜侧收集100 μL溶液,以通过以下公式一式一式两份定量粘膜FD4表观渗透系数(Papp)[24]:
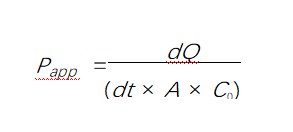
其中 dQ/dt 是传输速率(单位为 μg/s),对应于三个测量值的线性斜率,C0是供体室中的初始浓度,A是滑块的面积(0.71 cm 2)。
2.8. 通过 ELISA 测量炎症标志物
将冷冻空肠和回肠样品在液氮下粉碎,然后将100mg冷冻组织匀浆在1mL冷冻缓冲液中匀浆(货号89901,Thermo Fisher Scientific,美国马萨诸塞州沃尔瑟姆),然后在4°C下以10,000×g离心15分钟。收集上酯并在-20 °C下冷冻,用于随后的白细胞介素-1β(IL-1β),白细胞介素-8(IL-8)和肿瘤坏死因子-α(TNF-α)浓度的ELISA,根据制造商的说明使用商业试剂盒(货号DY681,DY535和DY690B,R&D Systems,明尼阿波利斯,密歇根州,美国)。
2.9. 甜菜碱分布
在血浆、尿液、肝脏、肾脏、肌肉、空肠、回肠和结肠组织中定量甜菜碱浓度。血浆浓度通过高压液相色谱法(HPLC)测定,如[25]所述,修改如[14]。首先将组织样品快速冷冻,然后在液氮中粉碎。然后将100mg粉碎的组织在1mL冷RIS缓冲液(1M,pH 7.0)中使用珠打浆器(Mini-BeadBeater-8,BioSpec产品,美国俄克拉荷马州巴特尔斯维尔)匀浆1分钟。然后将样品在4°C下以14,400×g离心20分钟。然后收集上酸盐,根据血浆样品进行分析,结果表示为mg 甜菜碱/ g组织。
2.10. 测试日粮血根碱浓度
用研钵和研杵压碎1克毛毡,在15mL的95:5乙腈:水中超声提取30分钟,然后以10,000×g离心10分钟。收集上消化液与10mL正己烷混合,然后以10,000×g离心3分钟。除去上清液并在30°C的氮气下干燥。将上消化液的残余物复溶于5 mL甲醇中,并向沉淀中加入15 mL 1%盐酸-甲醇(10:90 v / v),并在用5 mL甲醇复溶之前重复上述过程。将所得甲醇溶液混合并以12,000×g离心10分钟,然后通过0.22mm过滤器过滤,然后使用血根碱氯化物(>98%Sigma Aldrich,圣路易斯,密苏里州,美国)通过LCMS测定[26]所述。添加日粮中血根碱的浓度为0.61毫克/千克饲料。
2.11. 统计分析
该实验包括 2 × 3 因子设计,因此使用不平衡方差分析分析温度(热中性(TN) vs. 热应激(HS))、日粮(对照组(CON)、甜菜碱(BET)、异喹啉类生物碱(IQA))的合并主效应及其各自相互作用的参数(Genstat v. 18,VSN International,Hemel Hempstead,英国)。在适当的情况下,将时间包括在模型中,并使用实验重复(n = 5)作为阻塞因子。当存在偏斜数据时,例如尿甜菜碱和唾液皮质醇的情况,通过在Log10变换后重复分析来确认方差分析的有效性,从而降低了方差的异质性。文中提供的值是温度 (T)、日粮 (D) 或时间的合并主效应的校正均值和差值标准误差 (SED),而图表中显示的值是完全交互作用(T × D、T × D ×时间)。方差分析组在p ≤ 0.05时被认为存在显着差异,而在p ≤ 0.10时考虑趋势。事后费舍尔最小显着差异检验用于比较多个均值,使用字母上标在p < 0.05 处识别不同的组。
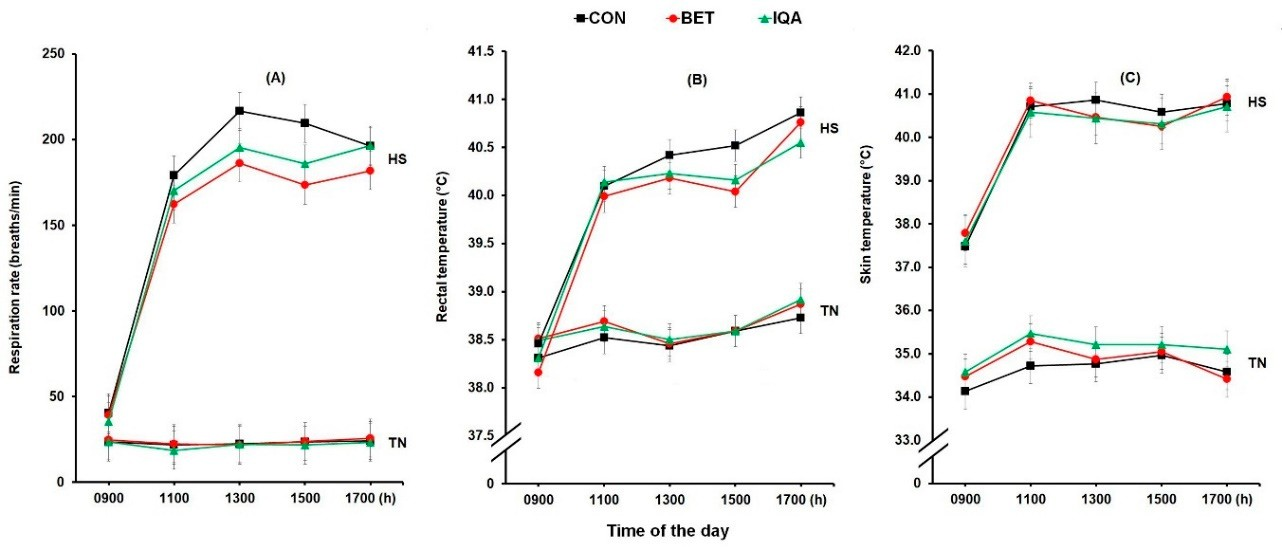
图1.在热中性(TN)或热应激条件(HS)下饲喂对照日粮(CON)、甜菜碱(BET)或异喹啉生物碱(IQA)日粮的生长猪日唿吸速率(A)、直肠温度(B)和皮肤温度(C)的变化。每个数字表示两天观测值的调整均值±合并的 SED × ×时间之间的相互作用。图(A)呼吸速率显着增加热应激(HS)(p < 0.001),一天中的时间(p < 0.001),甜菜碱(BET)和异喹啉类生物碱(IQA)低于整体对照组(CON)(日粮p = 0.024)和热应激(HS)期间(日粮*温度p = 0.013)。(B)热应激(HS)(p < 0.001)、一天中的时间(p < 0.001)显着升高直肠温度,甜菜碱(BET)和异喹啉类生物碱(IQA)低于热应激(HS)期间对照组(CON)组(日粮p = 0.001),而未观察到日粮的主要影响。(C)皮肤温度升高热应激(HS)(p < 0.001),一天中的时间(p < 0.001),并且在热应激(HS)期间异喹啉类生物碱(IQA)低于对照组(CON)(日粮p = 0.046),而没有观察到日粮的主要影响。温度×时间(p < 0.001)与呼吸速率、直肠温度和皮肤温度之间存在交互作用,而没有观察到温度×日粮×时间,每组代表n = 8-9头猪。
3. 结果
3.1. 生理反应
呼吸速率随着热应激HS介入而增加(23 vs 159次呼吸/分钟,p < 0.001),并随一天中的一段时间增加(在0900、1100、1300、1500和1700小时分别为31a、98b、109c、110c和113c唿吸/分钟;p < 0.001;图1A)。环境条件与时间之间存在相互作用(p < 0.001),使得与热中性TN条件下的猪相比,受到热应激HS的猪的呼吸速率RR在一天中呈增加趋势(图1A)。日粮治疗对呼吸速率RR有显着影响,因此饲喂甜菜碱BET或异喹啉类生物碱IQA的猪的呼吸速率RR均低于对照组CON(CON,BET和IQA分别为98a,88b和91b呼吸/分钟;p = 0.024)。此外,热应激HS与日粮处理之间也存在显着相互作用(p = 0.013),使得饲喂甜菜碱BET或异喹啉类生物碱IQA并接受热应激HS处理的猪的呼吸速率RR得到改善(数值较低),但在热中性TN条件下饲养的猪中则不然(图1A)。
直肠温度在900、1100、1300、1500和1700小时分别升高热应激(HS)(38.6 vs 39.9 ?C,p < 0.001)和一天中的时间(分别为38.4a、39.3b、39.4b、39.4b、39.4b和39.8c °C;p < 0.001;图 1B)。然而,环境条件和时间之间存在相互作用(p < 0.001),使得暴露于热应激(HS)的猪的直肠温度(RT)与接受热中性(TN)条件的猪相比更大程度地增加(图1B)。虽然日粮治疗对直肠温度(RT)没有主要影响,但热应激(HS)与日粮之间存在相互作用(p = 0.001),使得饲喂甜菜碱(BET)或异喹啉类生物碱(IQA)的猪的直肠温度(RT)较低并暴露于热应激(HS),但在热中性(TN)条件下的猪中则不然(图1B)。
对皮肤温度也观察到类似的影响。 皮肤温度在0900、1100、1300、1500和1700小时分别升高热应激(HS)(34.9 vs. 40.0 °C,p < 0.001)和一天中的时间(分别为36.0a、37.7b、37.8b、37.8b和37.9b °C;p < 0.001;图 1C)。然而,环境条件与发热时间之间存在交互作用(p < 0.001),使得受到热应激(HS)影响的猪在1100、1300、1500和1700 h时的皮肤温度(RT)高于0900 h,而其他时间则没有差异。虽然没有观察到日粮处理对皮肤温度(RT)的主要影响,但温度和日粮之间存在相互作用(p = 0.046),因此在热中性(TN)条件下,饲喂异喹啉类生物碱(IQA)的猪的皮肤温度(RT)高于对照组(CON)组,而所有其他组没有差异。
3.2. 血液血氧饱和度和电解质
血液pH值在热应激(HS)下降低(7.45 vs 7.43,p = 0.013,表2),但随着日粮治疗和环境条件与日粮之间的相互作用而保持不变。同样,对热应激(HS)的反应,血液部分氧(PO2)减少(67.2 vs 46.6 mmHg,p < 0.001)。日粮对PO2没有主要影响或交互作用。与对照组(CON)相比,血液O2饱和度降低(93.3 vs 82.0%,p < 0.001),异喹啉类生物碱(IQA)倾向于降低(对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA)分别为89.3%,88.8和86.2%;p = 0.096)。然而,环境条件与日粮之间存在相互作用(p = 0.035),因此饲喂异喹啉类生物碱(IQA)的猪的血O2饱和度低于饲喂的猪热应激(HS) 下的 对照组(CON) 和 甜菜碱(BET)(对照组(CON)、甜菜碱(BET) 和 异喹啉类生物碱(IQA) 分别为 85.2%、81.9% 和 78.2%),但在 热中性(TN) 条件下则不然。虽然二氧化碳分血压(PCO2)不受热应激(HS)的影响,但与对照组(CON)和异喹啉类生物碱(IQA)相比,甜菜碱(BET)倾向于降低该参数(对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA)分别为48.6,46.5和48.7mmHg;p = 0.096)。热应激倾向于降低总血二氧化碳(TCO2,34.8 vs. 33.6 mM,p = 0.074)和碳酸氢盐(HCO3,33.3 vs. 32.1 mM,p = 0.054)的浓度,而没有日粮或交互作用。血乳酸对 热应激(HS) 的反应降低(1.89 vs 1.15 mM,p = 0.001),但日粮处理没有主要或相互作用。血液中碱过量(BEb)降低热应激(HS)(8.16 vs. 6.97 mM,p = 0.029)。
虽然日粮处理没有主要效果,但有迹象表明环境条件与日粮之间存在相互作用(p = 0.088),使得饲喂甜菜碱(BET)或异喹啉类生物碱(IQA)的猪BEb在热应激(HS)期间与对照组(CON)相比下降幅度更大(对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA)分别为8.29,6.23和6.38mM)。对细胞外液中碱过量也有类似的效果(BEecf,表2)。细胞外液中碱过量响应热应激(HS)而减少(9.36 vs 8.01 mM,p = 0.032)。虽然日粮没有主要影响,但有迹象表明环境条件与日粮之间存在相互作用(p = 0.098),使得饲喂甜菜碱(BET)或异喹啉类生物碱(IQA)的猪的BEecf在热应激(HS)条件下比对照组(CON)猪的BEecf下降幅度更大(对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA)分别为9.54,7.14和7.35mM)。
血钾浓度响应热应激(HS)而升高(4.05 vs 4.30 mM,p = 0.001),但没有观察到日粮或相互作用的主要影响。同样,血氯化物浓度增加热应激(HS)(98.4 vs 100.4 mM,p = 0.027),但没有日粮或相互作用的主要影响。环境条件或日粮治疗对钠、钙、阴离子间隙和葡萄糖的血液浓度没有影响(表2)。热应激降低了血细胞比容(35.3 vs 31.8%,p < 0.001)和血红蛋白浓度(11.9 vs 10.8 g/dL,p < 0.001),但对这些参数没有日粮或交互影响(表2)。
表 2.热中性 (TN) 和热应激 (HS) 条件和对照组(CON)、甜菜碱 (BET) 或异喹啉生物碱 (IQA) 日粮对血气和电解质的影响 1.
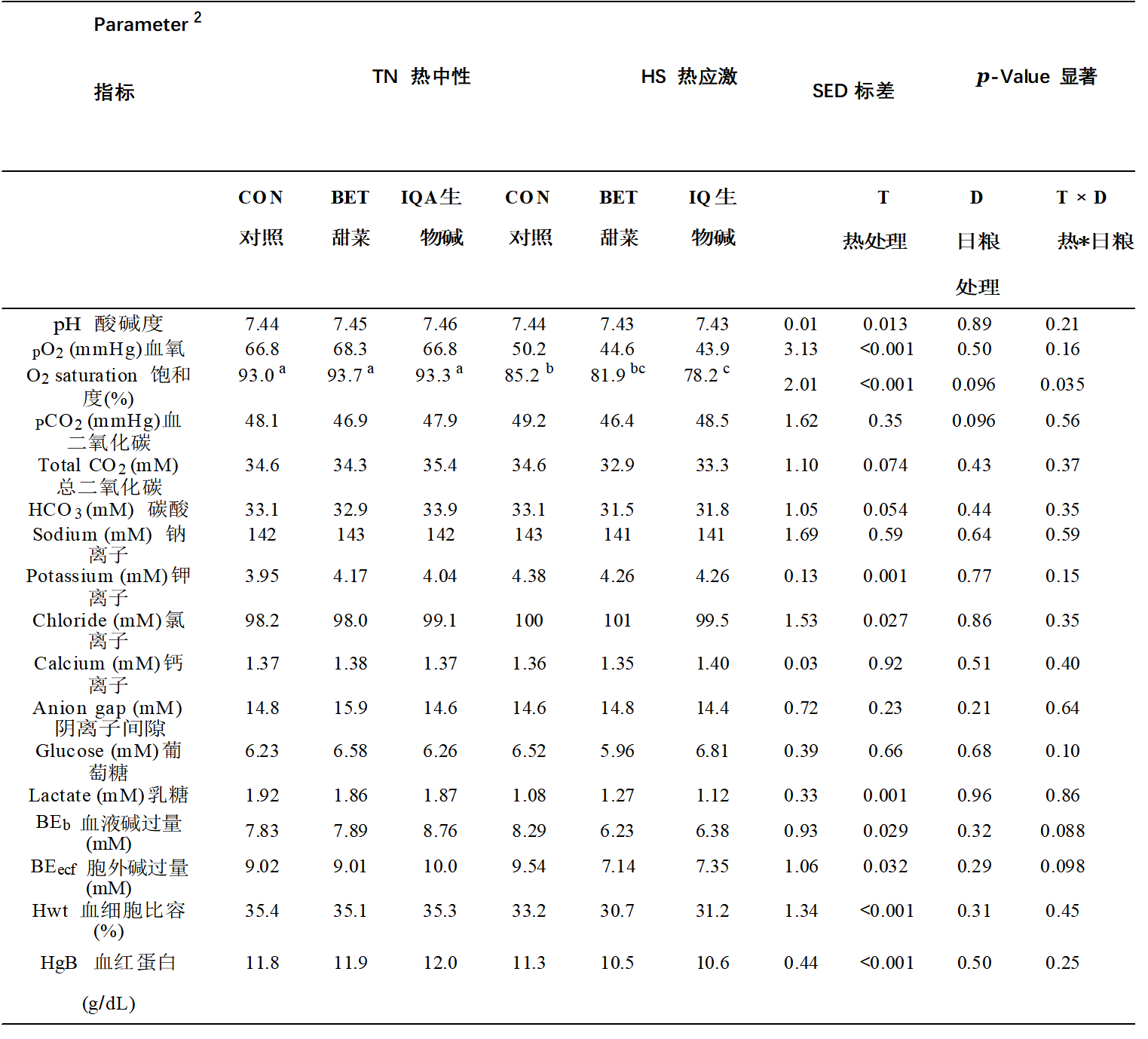
1 n = 每组 8-9 头猪,值代表 T D 之间相互作用的调整均值合并标准差值 (SED)。 2 BEb:血液中碱过量;BEecf:额外细胞液中的碱过量;Hwt:血细胞比容;HgB:血红蛋白。a–c 不同的上标表示组 p < 0.05。
3.3. 血液和尿液生化
血浆肌酐响应热应激(HS)而升高(0.126 vs 0.146 mM,p < 0.001,表3),但不受日粮治疗的影响。热应激对血浆白蛋白浓度没有影响,但日粮有一个主要影响,即甜菜碱(BET)和异喹啉类生物碱(IQA)日粮中的猪白蛋白浓度低于对照组(CON)中的白蛋白浓度(对照组(CON)、甜菜碱(BET)和异喹啉类生物碱(IQA)分别为44.4、39.9和40.1 g/L;p = 0.048)。环境条件和日粮处理或相互作用对血浆蛋白、胆红素和甘油三酯浓度没有主要影响(表3)。
表 3.热中性 (TN) 和热应激 (HS) 条件和控制 (CON)、甜菜碱 (BET) 或异喹啉生物碱 (IQA) 日粮对血浆生化和激素的影响 1.
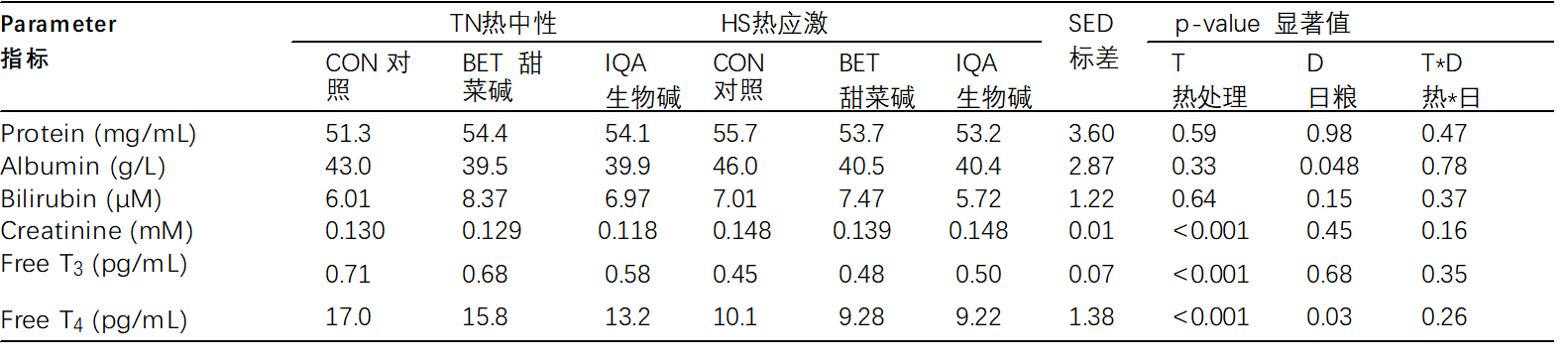
1 n = 每组 8-9 头猪,值表示调整平均值 ± T × D 之间相互作用的合并标准差值 (SED)。 a–c 不同的上标表示组 p < 0.05。
表 4.热中性(TN)和热应激(HS)条件和对照组(CON),甜菜碱(BET)或异喹啉生物碱(IQA)日粮对尿液生化和代谢物的影响 1.
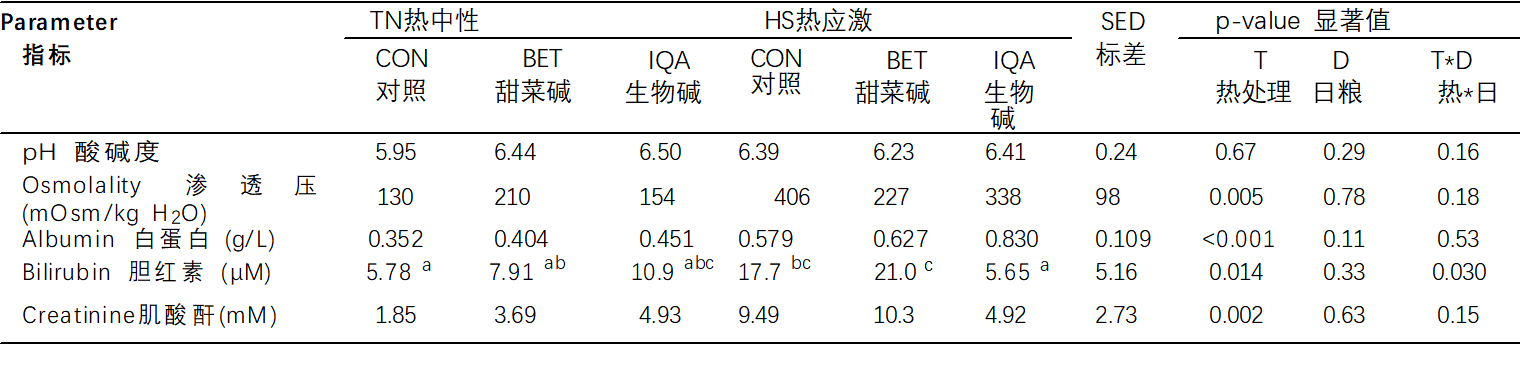
1 n = 每组 8-9 头猪,值表示调整平均值 ± T × D 之间相互作用的合并标准差值 (SED)。 a–c 不同的上标表示组 p < 0.05。
3.4. 热应激(HS)反应对内分泌的影响
唾液皮质醇在 热应激(HS) 后增加(25.2 vs 54.6 ng/mL,p = 0.010,图 2)。日粮治疗对唾液皮质醇没有主要(p = 0.56)或相互作用(p = 0.75)的影响。无血浆T3响应热应激(HS)而降低(0.66 vs 0.47 pg/mL,p < 0.001,表3),但没有日粮或相互作用的主要影响。 同样,无血浆T4响应热应激(HS)而降低(15.4 vs. 9.55 pg/mL,p < 0.001,表 3)。日粮处理对血浆游离T4有显着影响,因此饲喂异喹啉类生物碱(IQA)的猪的该参数低于对照组(CON)和甜菜碱(BET)的猪(对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA)分别为13.6a,12.5a和11.2b ng / dL;p = 0.033,表3)。环境条件与日粮对血浆游离T4无交互作用。
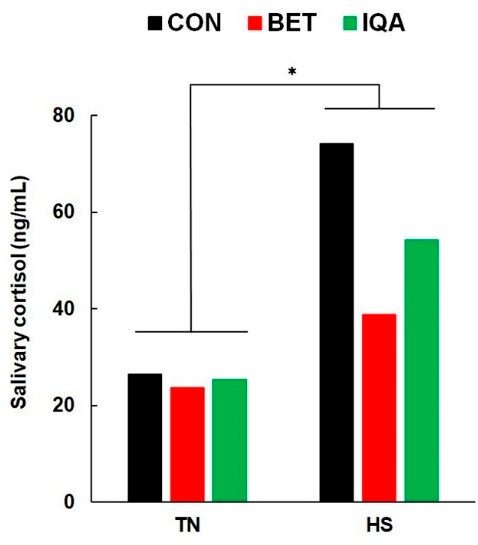
图2.热中性 (TN) 和热应激 (HS) 条件对饲喂对照日粮 (CON)、对照加甜菜碱 (BET) 或对照加异喹啉生物碱 (IQA) 的生长猪唾液皮质醇浓度的影响。每个条形代表在热挑战的第 1 天和第 2 天收集的唾液的汇集效果。由于样品异质性,在分析前对结果进行变换(log10),然后表示为反向变换均值的平均值。热应激(HS)显着增加唾液皮质醇(* p = 0.010)。未观察到日粮或与温度相互作用的影响。n = 每组 8-9 头猪,值表示 T × D 之间相互作用的合并 SED ±调整均值。
3.5. 炎症和抗氧化标志物
白细胞介素-1β浓度在空肠(125 vs 193 pg/mL,p = 0.002,表5)和回肠(299 vs 414 pg/mL,p = 0.004)中响应热应激(HS)而增加。虽然日粮没有主要影响,但有迹象表明环境条件和日粮对空肠IL-1β有相互作用(p= 0.056),使得与对照组(CON)和异喹啉类生物碱(IQA)猪相比,甜菜碱(BET)上的热应激(HS)猪倾向于增加IL-1β浓度(对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA)分别为173,253和154 pg/mL)。回肠中的 热应激(HS) 增加了白细胞介素-8 浓度(2639 vs 3108 pg/mL,p = 0.012),但在空肠中保持不变。对IL-8没有主要或互动的日粮影响。热应激(HS)或日粮对TNF-α没有影响(表5)。红细胞沉降率(ESR)通过热应激(HS)增加(9.4 vs. 12.5 mm/h,p = 0.021,表5),但日粮不变。环境条件对血浆TAC没有主要影响。然而,日粮治疗有显着效果,使得TAC在甜菜碱(BET)中高于异喹啉类生物碱(IQA),但两种日粮与对照组(CON)都没有显着差异(分别为1.36a,1.62ab,1.19ac mM Trolox用于对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA);p = 0.020,表5)。
表 5.热中性(TN)和热应激(HS)条件和对照组(CON),甜菜碱(BET)或异喹啉生物碱(IQA)对肠道炎症反应的影响1
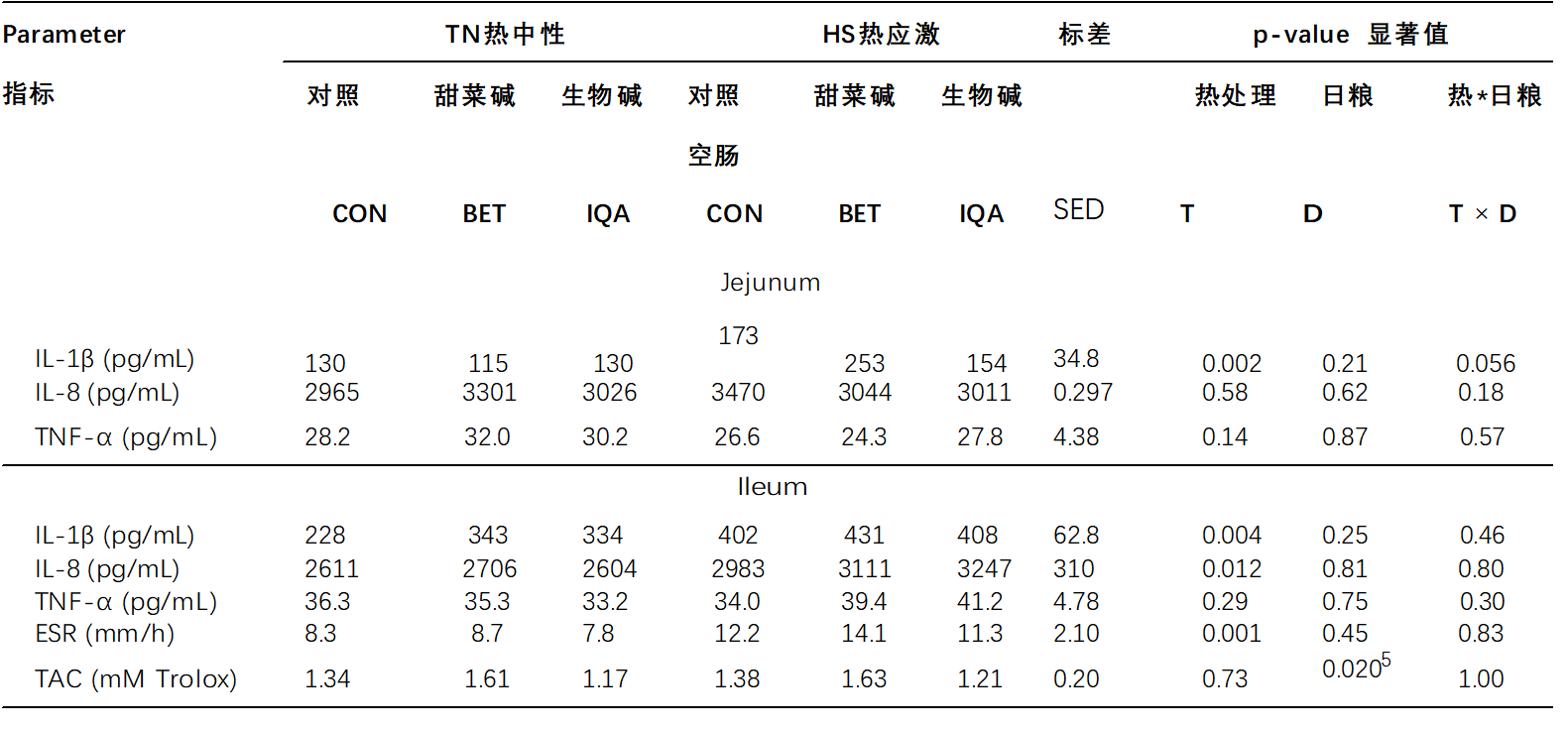
1 n = 每组 8-9 头猪,值表示 T × D 之间相互作用的调整均值±合并标准差值 (SED)。
3.6. 肠道通透性
回肠通透性不受温度或日粮的影响(图3A)。虽然热应激(HS)或日粮对结肠通透性没有主要影响,但温度和日粮之间存在显着的相互作用(p = 0.030),使得在热应激(HS)条件下饲喂异喹啉类生物碱(IQA)的猪的FD4渗透性低于对照组(CON)(对照组(CON),甜菜碱(BET)和异喹啉类生物碱(IQA)分别为1566,1040和756×10-4cm/sec),而所有其他组不显着(图3B)。
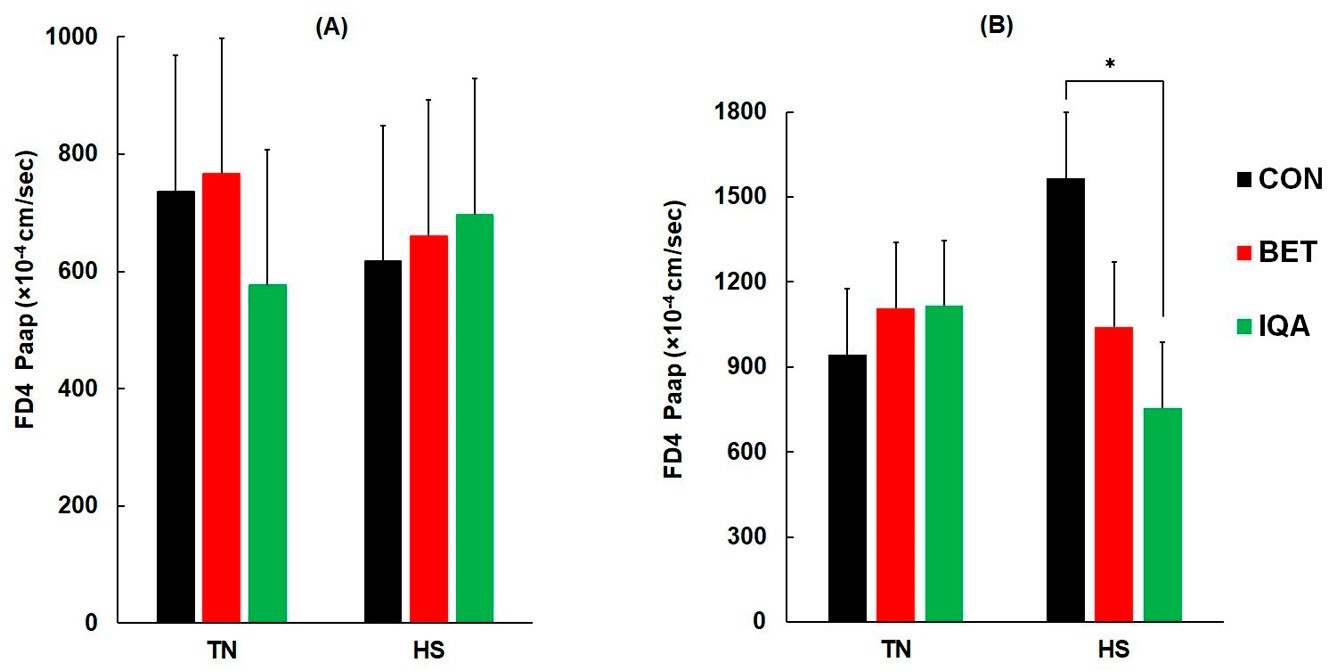
图3.热应激(HS)和热中性(TN)条件对回肠(A)和结肠的影响
(B)异硫氰酸荧光素右旋糖酐(4 k Da)对饲喂对照日粮(CON)、对照加甜菜碱(BET)或对照加异喹啉生物碱(IQA)的生长猪的表观渗透系数(FD4 Papp)。结果调整平均值±SED和n = 8-9头猪。(A)温度、日粮和日粮*温度影响的p值分别为0.77、0.90和0.71。(B)温度、日粮和日粮*温度影响的p值分别为0.56、0.23和0.030。* 表示由费舍尔最小显着差异检验确定的 p < 0.05。
3.7. 甜菜碱处理分析
添加甜菜碱增加了血浆中(对照组(CON)、甜菜碱(BET)和异喹啉类生物碱(IQA)分别为840a、1282b和811a μmol/L)的甜菜碱浓度,p<0.001,表6)、尿液(1910a、5957b和1570a μmol/L,p = 0.002)、肝脏(1.16a、1.55b和1.18a mg/g,p = 0.007)、肾脏(1.00a、1.12b和0.71a mg/g p<0.001)、空肠(0.37a、0.49b和0.35a和mg/g p<0.001)和回肠(0.47a、0.58b和0.44a mg/g p < 0.001)甜菜碱浓度也均有上升。甜菜碱(BET)处理组在肌肉中的甜菜碱浓度往往较高(0.52,0.74和0.58mg / g,p = 0.080),但没有观察到结肠甜菜碱浓度增加。无论日粮如何,热应激(HS)猪的甜菜碱浓度在血浆中(923 vs. 1021 μmol/L, p = 0.08) 和肝脏(1.20 vs. 1.38 mg/g, p = 0.07)普遍更高。在热中性(TN)条件下(0.45,0.61和0.48 mg/g,相互作用p = 0.097),甜菜碱(BET)倾向于增加回肠甜菜碱浓度,但在热应激(HS)中未观察到趋势。
表 6.热中性(TN)和热应激(HS)条件和对照组(CON)、甜菜碱(BET)或异喹啉生物碱(IQA)对生长猪甜菜碱分布的影响 1.
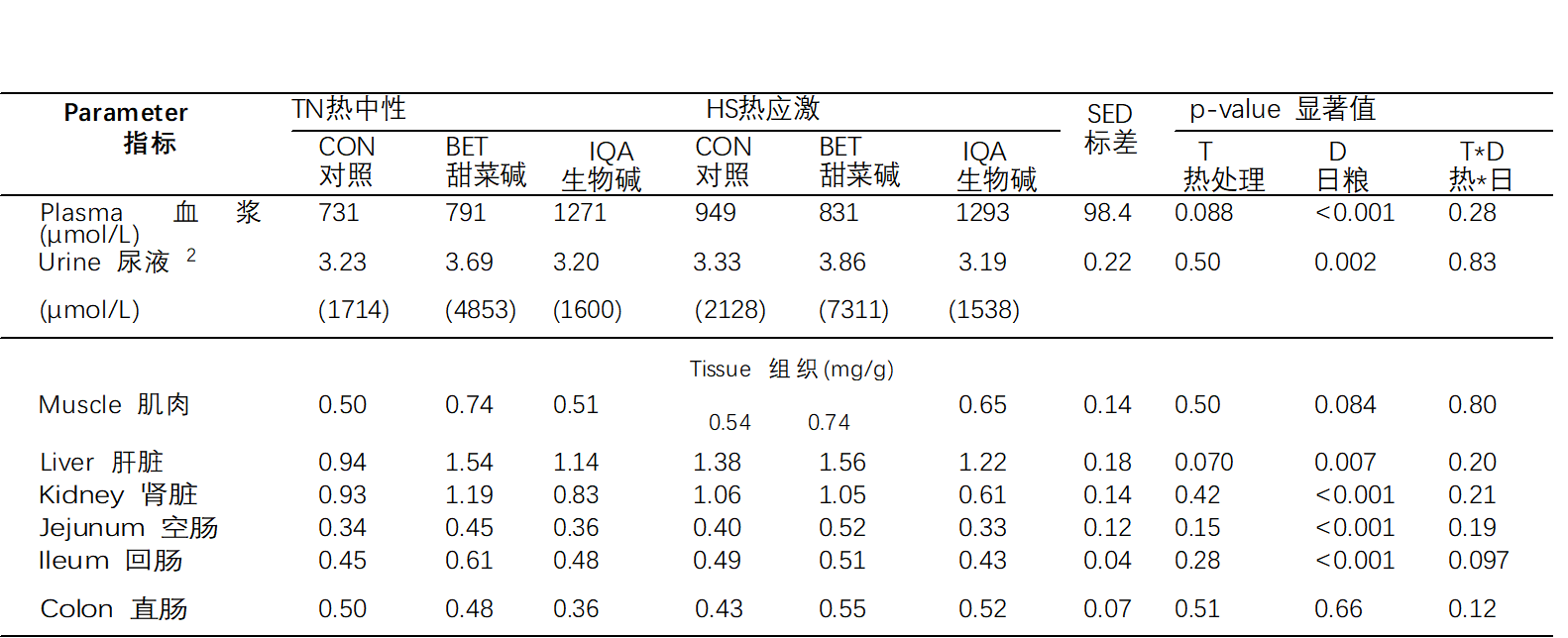
1 n = 每组 8-9 头猪,值代表调整均值 T D 之间交互作用的合并差值标准误差 (SED)。 2 数据偏斜,因此在分析和反向变换均值之前进行 log10 变换在括号中。
4. 讨论
本研究的主要发现是,添加甜菜碱和异喹啉生物碱部分改善了生长猪对热应激(HS)的体温调节反应。呼吸速率(RR)降低与直肠温度(RT)降低的组合证明了这一点。这一发现的意义在于,由于猪缺乏产生汗腺的水[27],因此它们在热应激(HS)期间依靠“蒸发喘气”来增加对环境的热量损失。在这个过程中,流向上呼吸道的血流量增加,呼吸模式改变,潮气量减少,但呼吸频率[28]和流向上呼吸道的血流量增加[4]。这增加了上呼吸道的气流,通过流口水的蒸发实现冷却。因此,这些结果表明,甜菜碱(BET)和异喹啉类生物碱(IQA)日粮通过降低核心体温(RT)以及降低温度所需的努力来改善热应激(HS),正如较低的呼吸速率(RR)所证明的那样。
高热期间肠血流量减少可损害胃肠道(GIT)血流,导致上皮细胞丢失和固有层暴露,这在回肠中与TER降低以及对大分子和细菌脂多糖的通透性增加有关[8,29,30]。与已发表的数据一致,我们发现与热中性(TN)猪相比,热应激(HS)的结肠大分子通透性高出约60%。结肠对营养调整有反应,异喹啉类生物碱(IQA)阻止了热应激(HS)猪中发生的大分子通透性增加,表明粘膜完整性得到改善。血根碱对减少结肠渗漏的有益作用已在大鼠结肠炎模型的其他地方得到证实[31],这些结果共同支持结肠是异喹啉类生物碱(IQA)的作用部位。使用的异喹啉类生物碱(IQA)配方来源于Macleaya cordata,也称为“羽状罂粟”,并且富含异喹啉类生物碱(IQA),如血根碱[21]。Macleaya cordata提取物已被证明可以改善胃肠道(GIT)健康,如降低腹泻评分和沙门氏菌种群,同时促进黏膜生长[17,20,32]。在另一项实验中,博落回Macleaya cordata提取物减少了沙门氏菌,增加了乳酸菌的数量,抗氧化能力,免疫功能参数,小肠绒毛高度和生长性能[20]。
异喹啉类生物碱(IQA)似乎有多种机制可以使胃肠道(GIT)粘膜受益。这包括抗菌和驱虫特性[33]。此外,其他绵羊实验观察到Macleaya cordata提取物可减少氧化应激[34],包括在热应激动物中观察到饲料转换效率的提高[35]。据广泛报道,异喹啉生物碱具有抗炎特性[31,36],其作用至少部分通过NF-kβ的变化介导[37]。虽然本实验中使用的热应激(HS)模型增加了小肠和血液ESR的炎症,血ESR也被用作炎症的标志物,但没有观察到异喹啉类生物碱(IQA)对炎症标志物的影响。值得注意的是,血根碱的生物利用度较差[38-40],但猪微生物组和肠黏膜的代谢已被量化[40]。在该实验中,结肠粘膜是血根碱代谢为二羟基血根碱(DHSA)的最大部位,而回肠微生物组是微生物组代谢最活跃的区域。这将胃肠道(GIT)的远端区域定位为血根碱代谢的最重要部位,这与当前实验中结肠是通透性降低的部位一致。
由于血根碱等异喹啉类生物碱(IQA)的生物利用度低,唿吸速率(RR)和直肠温度(RT)的降低是出乎意料的。在单次口服剂量为10mg/kg体重的血根碱中,只有15%的血根碱通过尿液排泄,但17.3%的剂量保留在肝脏中,29%保留在胃肠道(GIT)中,3.6%保留在肾脏中[41],表明一些血根碱具有生物可利用性,因此可能影响胃肠道(GIT)以外的体温调节。胃肠道(GIT)内可能存在有助于减少产热的重要作用,即对Na+/K+ATP酶泵活性的抑制作用[18,42]。这种泵活动对产热有很大贡献,提供了高达72%的动物产热[19]。甜菜碱还通过降低Na+/K+ATP酶活性来减少产热[10],尽管甜菜碱的生物利用度要高得多。此外,异喹啉类生物碱(IQA)可以减少大肠中的发酵,这也有助于产生热量[43]。在其他地方,日粮异喹啉类生物碱(IQA)已被证明具有吸收后作用,例如抗氧化能力的改善,并且还可以降低运输应激后的皮质醇浓度[16],支持胃肠道(GIT)以外的效果。考虑到这些结果与[38]相比,[38]表明只有一小部分生物碱被吸收,血根碱和其他异喹啉生物碱在胃肠道(GIT)之外的影响值得进一步研究。
与异喹啉生物碱一样,添加甜菜碱可改善热应激(HS)生长猪呼吸速率(RR)和直肠温度(RT)的增加,密切反映了肉鸡[11,14]和绵羊[44]的结果。甜菜碱至少具有三种已知的生物学功能,作为甲基供体、渗透剂或直接作为协同[9]。甜菜碱的代谢主要是作为蛋氨酸循环中同型半胱氨酸的甲基供体。该反应由甜菜碱同型半胱氨酸甲基转移酶(BHMT)催化,BHMT主要在肝脏中表达,在肾脏中表达程度较小[45]。添加甜菜碱在空肠和回肠中增加,但结肠没有增加,这支持研究表明甜菜碱具有高生物利用度[46,47]。然而,尽管空肠和回肠升高,但未观察到黏膜Papp的改善。这与[48]的结果不同,后者观察到回肠通透性降低。重要的是,[48]的研究使用了0.125%甜菜碱的剂量,而不是当前研究中的0.1%,并支持使用更高剂量的甜菜碱来改善热应激(HS)。观察到添加甜菜碱在肝脏和肾脏中的分布增加,这与其他研究一致[14,46],这些器官是甜菜碱利用的关键部位。在人类中,甜菜碱的主要代谢命运是代谢为二甲基甘氨酸[47]。值得注意的是,在目前的实验中,未修饰的甜菜碱似乎有显着的排泄,这表明当前实验中甜菜碱的一部分作用是由于直接渗透作用,例如降低Na+/K+-ATP酶和Ca2+-ATP酶的活性[10]或协同效应。
5. 结论
异喹啉类生物碱(IQA)和甜菜碱(BET)都改善了生长猪的热应激,但似乎通过不同的机制来实现。异喹啉生物碱似乎主要作用于胃肠道(GIT), 提供了防止热应激引起的结肠通透性增加的额外好处.相比之下,甜菜碱似乎很容易从胃肠道(GIT)吸收,结肠之前部位从胃肠道(GIT)组织中消失耗竭。未修饰的甜菜碱的尿排泄表明部分作用与其是细胞渗透剂有关,但也观察到对作为甜菜碱代谢部位的肝脏和肾脏的分布增加。

-
[06-25]
-
[06-15]
-
[05-22]
-
[05-12]
-
[04-23]
-
[04-14]
-
[04-07]
-
[02-18]
-
[02-03]
-
[01-13]


